
Do mesmo modo que uma fábrica pode ser multiplicada pela construção de várias filiais, também as células se dividem e produzem cópias de si mesmas.
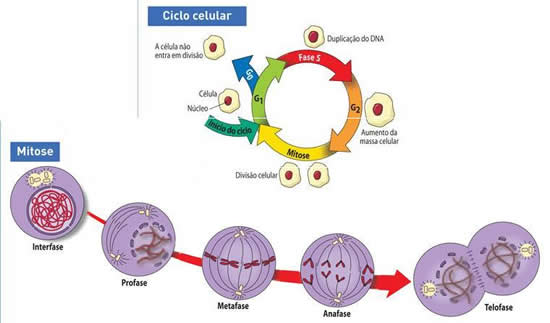
Há dois tipos de divisão celular: mitose e meiose.
Na mitose, a divisão de uma “célula-mãe” duas “células-filhas” geneticamente idênticas e com o mesmo número cromossômico que existia na célula-mãe. Uma célula n produz duas células n, uma célula 2n produz duas células 2n etc. Trata-se de uma divisão equacional.
Já na meiose, a divisão de uma “célula-mãe” 2n gera “células-filhas” n, geneticamente diferentes. Neste caso, como uma célula 2n produz quatro células n, a divisão é chamada reducional.
A interfase – A fase que precede a mitose
É impossível imaginar a multiplicação de uma fabrica, de modo que todas as filiais fossem extremamente semelhantes a matriz, com cópias fieis de todos os componentes, inclusive dos diretores? Essa, porém, no caso da maioria das células, é um acontecimento rotineiro. A mitose corresponde a criação de uma cópia da fabrica e sua meta é a duplicação de todos os componentes.A principal atividade da célula, antes de se dividir, refere-se a duplicação de seus arquivos de comando, ou seja, à reprodução de uma cópia fiel dos dirigentes que se encontram no núcleo.
A interfase é o período que precede qualquer divisão celular, sendo de intensa atividade metabólica.Nesse período, há a preparação para a divisão celular, que envolve a duplicação da cromatina, material responsável pelo controle da atividade da célula. Todas as informações existentes ao longo da molécula de DNA são passadas para a cópia, como se correspondessem a uma cópia fotográfica da molécula original. Em pouco tempo, cada célula formada da divisão receberá uma cópia exata de cada cromossomo da célula se dividiu.
As duas cópias de cada cromossomo permanecem juntas por certo tempo, unidas pelo centrômero comum, constituindo duas cromátides de um mesmo cromossomo. Na interfase, os centríolos também se duplicam.
A interfase e a Duplicação do DNA
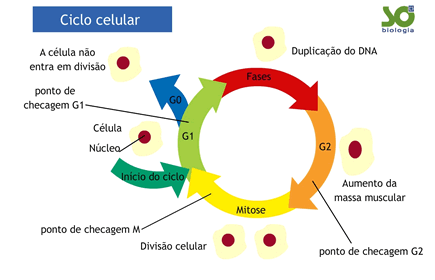
Houve época em que se falava que a interfase era o período de “repouso” da célula. Hoje, sabemos, que na realidade a interfase é um período de intensa atividade metabólica no ciclo celular: é nela que se dá aduplicação do DNA, crescimento e síntese. Costuma-se dividir a interfase em três períodos distintos:G1, S e G2.O intervalo de tempo em que ocorre a duplicação do DNA foi denominado de S (síntese) e o período que antecede é conhecido como G1 (G1 provém do inglês gap, que significa “intervalo”). O período que sucede o S é conhecido como G2.
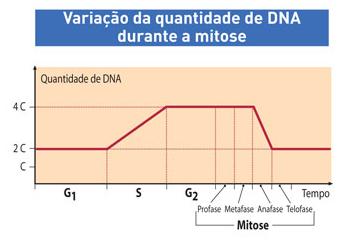
O ciclo celular todo, incluindo a interfase (G1, S, G2) e a mitose (M) – prófase, metáfase, anáfase e telófase – pode ser representado em um gráfico no qual se coloca a a quantidade da DNA na ordenada (y) e o tempo na abscissa (x). Vamos supor que a célula que vai se dividir tenha, no período G1, uma quantidade 2C de DNA (C é uma unidade arbitrária). O gráfico da variação de DNA, então, seria semelhante ao da figura abaixo.
Nas células, existe uma espécie de “manual de verificação de erros” que é utilizado em algumas etapas do ciclo celular e que é relacionado aos pontos de checagem. Em cada ponto de checagem a célula avalia se é possível avançar ou se é necessário fazer algum ajuste, antes de atingir a fase seguinte. Muitas vezes, a escolha é simplesmente cancelar o processo ou até mesmo conduzir a célula à morte.
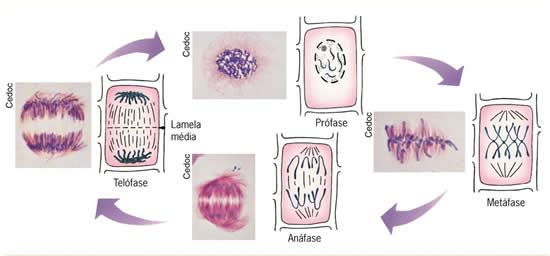
As fases da mitose
A mitose é um processo contínuo de divisão celular, mas, por motivos didáticos, para melhor compreendê-la, vamos dividi-la em fases: prófase, metáfase, anáfase e telófase. Alguns autores costumam citar uma quinta fase – a prometáfase – intermediária entre a prófase e a metáfase. O final da mitose, com a separação do citoplasma, é chamado de citocinese.Prófase – Fase de início (pro = antes)
Os cromossomos começam a ficar visíveis devido à espiralação.
O nucléolo começa a desaparecer.
Organiza-se em torno do núcleo um conjunto de fibras (nada mais são do que microtúbulos) originadas a partir dos centrossomos, constituindo o chamado fuso de divisão (ou fuso mitótico).
Embora os centríolos participem da divisão, não é deles que se originam as fibras do fuso. Na mitose em célula animal, as fibras que se situam ao redor de cada par de centríolos opostas ao fuso constituem o áster (do grego, aster = estrela).
O núcleo absorve água, aumenta de volume e a carioteca se desorganiza.
No final da prófase, curtas fibras do fuso, provenientes do centrossomos, unem-se aos centrômeros. Cada uma das cromátides-irmãs fica ligada a um dos pólos da célula.
Note que os centrossomos ainda estão alinhados na região equatorial da célula, o que faz alguns autores designarem essa fase de prometáfase.
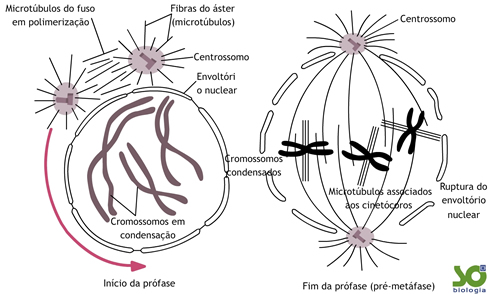
A formação de um novo par de centríolos é iniciada na fase G1, continua na fase S e na fase G2 a duplicação é completada. No entanto, os dois pares de centríolos permanecem reunidos no mesmo centrossomo. Ao iniciar a prófase, o centrossomo parte-se em dois e cada par de centríolos começa a dirigir-se para pólos opostos da célula que irá entrar em divisão.
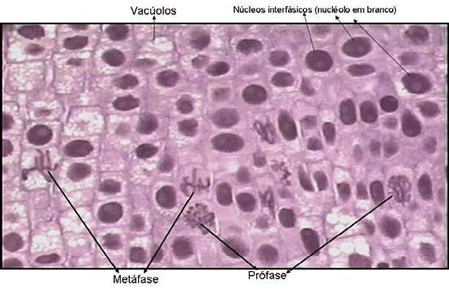
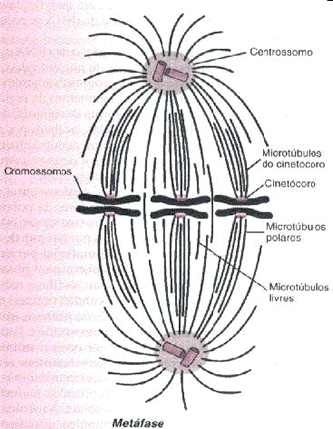
Metáfase – Fase do meio (meta = no meio)
Os cromossomos atingem o máximo em espiralação, encurtam e se localizam na região equatorial da célula.No finalzinho da metáfase e início da anáfase ocorre a duplicação dos centrômeros.
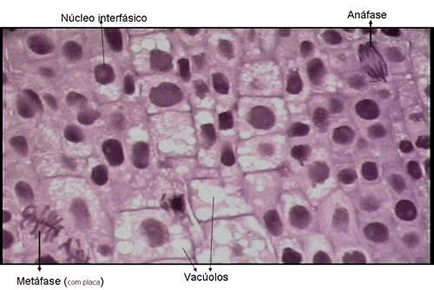
Anáfase – Fase do deslocamento (ana indica movimento ao contrário)
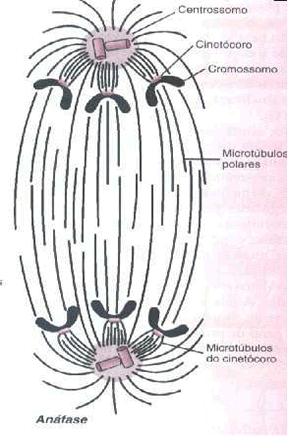
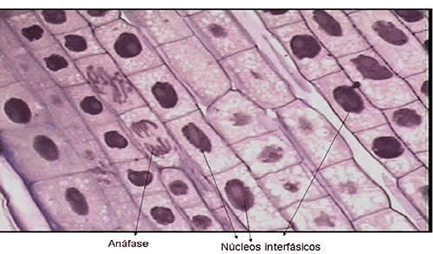
As fibras do fuso começam a encurtar. Em conseqüência, cada lote de cromossomos-irmãos é puxado para os pólos opostos da célula.
Como cada cromátide passa a ser um novo cromossomo, pode-se considerar que a célula fica temporariamente tetraplóide.
Telófase – Fase do Fim (telos = fim)
Os cromossomos iniciam o processo de desespirilação.Os nucléolos reaparecem nos novos núcleos celulares.
A carioteca se reorganiza em cada núcleo-filho.
Cada dupla de centríolos já se encontra no local definitivo nas futuras células-filhas.
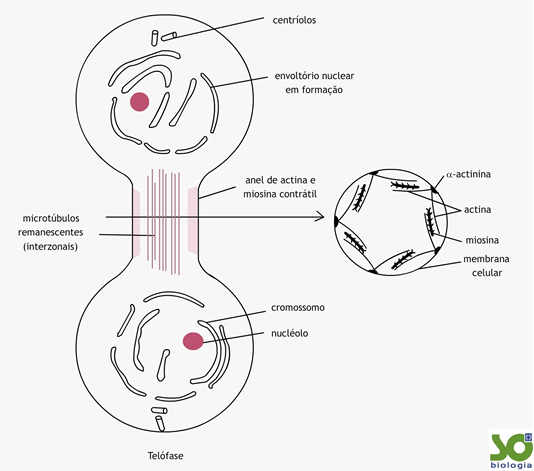
Citocinese – Separando as células
A partição em duas copias é chamada de citocinese e ocorre, na célula animal, de fora para dentro, isto é, como se a célula fosse estrangulada e partida em duas (citocinese centrípeta). Há uma distribuição de organelas pelas duas células-irmãs. Perceba que a citocinese é, na verdade a divisão do citoplasma. Essa divisão pode ter início já na anáfase, dependendo da célula.
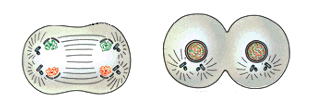
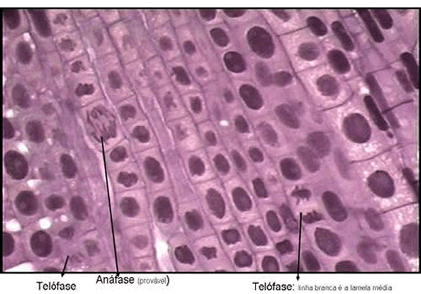
A Mitose na Célula Vegetal
Na mitose de células de vegetais superiores, basicamente duas diferenças podem ser destacadas, em comparação com que ocorre na mitose da célula animal:A mitose ocorre sem centríolos. A partir de certos locais, correspondentes ao centrossomos, irradiam-se as fibras do fuso. Uma vez que não há centríolos, então não existe áster. Por esse motivo, diz-se que a mitose em células vegetais é anastral (do grego, an = negativo);
A citocinese é centrífuga, ocorre do centro para a periferia da célula. No início da telófase forma-se o fragmoplasmo, um conjunto de microtúbulos protéicos semelhantes aos do fuso de divisão. Os microtúbulos do fragmoplasto funcionam como andaimes que orientam a deposição de uma placa celular mediana semelhante a um disco, originada de vesículas fundidas do sistema golgiense. Progressivamente, a placa celular cresce em direção à periferia e, ao mesmo tempo, no interior da vesícula, ocorre a deposição de algumas substâncias, entre elas, pectina e hemicelulose, ambos polissacarídeos. De cada lado da placa celular, as membranas fundidas contribuem para a formação, nessa região, das membranas plasmáticas das duas novas células e que acabam se conectando com a membrana plasmática da célula-mãe. Em continuação à formação dessa lamela média, cada célula-filha, deposita uma parede celulósica primária, do lado de fora da membrana plasmática. A parede primária acaba se estendendo por todo o perímetro da célula. Simultaneamente a parede celulósica primária da célula-mãe é progressivamente desfeita, o que permite o crescimento de cada célula-filha, cada qual dotada, agora, de uma nova parede primária. Então, se pudéssemos olhar essa região mediana de uma das células, do citoplasma para fora, veríamos, inicialmente, a membrana plasmática, em seguida a parede celulósica primária e, depois, a lamela média. Eventualmente, uma parede secundária poderá ser depositada entre a membrana plasmática e a parede primária.
A mitose serve para...
A mitose é um tipo de divisão muito freqüente entre os organismos da Terra atual. Nos unicelulares, serve à reprodução assexuada e à multiplicação dos organismos. Nos pluricelulares, ela repara tecidos lesados, repões células que normalmente morrem e também está envolvida no crescimento.
No homem, a pele, a medula óssea e o revestimento intestinal são locais onde a mitose é freqüente. Nem todas as células do homem, porém, são capazes de realizar mitose. Neurônios e célula musculares são dois tipos celulares altamente especializados em que não ocorre esse tipo de divisão (ocorre apenas na fase embrionária). Nos vegetais, a mitose ocorre em locais onde existem tecidos responsáveis pelo crescimento, por exemplo, na ponta de raízes, na ponta de caules e nas gemas laterais. Serve também para produzir gametas, ao contrário do que ocorre nos animais, em que a meiose é o processo de divisão mais diretamente associado à produção das células gaméticas.
O Controle do Ciclo Celular e a Origem do Câncer
Como sabemos, a interfase é um período de intensa atividade metabólica e de maior duração do ciclo celular. Células nervosas e musculares, que não se dividem por mitose, mantêm-se permanentemente na interfase, estacionadas no período chamado G0.Nas células que se dividem ativamente, a interfase é seguida da mitose, culminando na citocinese. Sabe-se que a passagem de uma fase para outra é controlada por fatores de regulação - de modo geral protéicos – que atuam nos chamados pontos de checagem do ciclo celular. Dentre essas proteínas, se destacam as ciclinas, que controlam a passagem da fase G1 para a fase S e da G2 para a mitose.
Se em algumas dessas fases houver alguma anomalia, por exemplo, algum dano no DNA, o ciclo é interrompido até que o defeito seja reparado e o ciclo celular possa continuar. Caso contrário, a célula é conduzida à apoptose (morte celular programada).
Outro ponto de checagem é o da mitose, promovendo a distribuição correta dos cromossomos pelas células-filhas. Perceba que o ciclo celular é perfeitamente regulado, está sob controle de diversos genes e o resultado é a produção e diferenciação das células componentes dos diferentes tecidos do organismo. Os pontos de checagem correspondem, assim, a mecanismos que impedem a formação de células anômalas.
A origem das células cancerosas está associada a anomalias na regulação do ciclo celular e à perda de controle da mitose. Alterações do funcionamento de genes controladores do ciclo celular, em decorrência de mutações, são relacionados ao surgimento de um câncer. Duas classes de genes, os proto-onco-genes e os genes supressores de tumor são os mais diretamente relacionados à regulação do ciclo celular. Os proto-oncogenes são responsáveis pela produção de proteínas que atuam na estimulação do ciclo celular, enquanto os genes supressores de tumor são responsáveis pela produção de proteínas que atuam inibindo o ciclo celular.
Dizendo de outro modo:
Os proto-oncogenes, quando ativos, estimulam a ocorrência de divisão celular e os genes supressores de tumor, quando ativos, inibem a ocorrência de divisão celular. O equilíbrio na atuação desses dois grupos de genes resulta no perfeito funcionamento do ciclo celular.
Mutações nos proto-oncogenes os transformam em oncogenes ( genes causadores de câncer). As que afetam os genes supressores de tumor perturbam o sistema inibidor e o ciclo celular fica desregulado, promovendo a ocorrência desordenada de divisões celulares e o surgimento de células cancerosas, que possuem as seguintes características:
são indiferenciadas, não contribuindo para a formação natural dos tecidos,
seus núcleos são volumosos e com um número anormal de cromossomos;
empilham-se sobre a outras em várias camadas, originando um aglomerado de células que forma um tumor. Se ficar restrito ao local de origem e for encapsulado, diz-se que o tumor é benigno, podendo ser removido;
nos tumores malignos, ocorre a metástase, ou seja, as células cancerosas abandonam o local de origem, espalham-se por via sangüínea ou linfática, e invadem outros órgãos. Esse processo é acompanhado por uma angiogênese, que é a formação de inúmeros vasos sanguíneos responsáveis pela nutrição das células cancerosas.
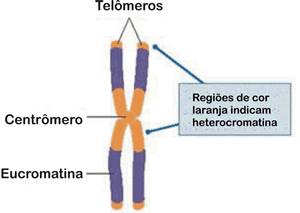
Outra ocorrência envolvendo alterações do ciclo celular é relativa aos telômeros, que são segmentos de moléculas de DNA com repetições de bases que atuam como “capas protetoras” da extremidade dos cromossomos.
Em células humanas normais, a cada ciclo celular os telômeros são progressivamente encurtados, as extremidades dos cromossomos ficam cada vez mais curtas, até atingir um limite mínimo de tamanho incompatível com a vida da célula, paralisando-se as divisões celulares e sinalizando o fim da vida da célula.
Em células cancerosas esse limite é transposto graças a atividade de uma enzima , atelomerase, que atua na reposição constante dos telômeros, mantendo-os sempre com o tamanho original, permitindo assim, que as células se dividam continuamente e se tornem praticamente “imortais”.
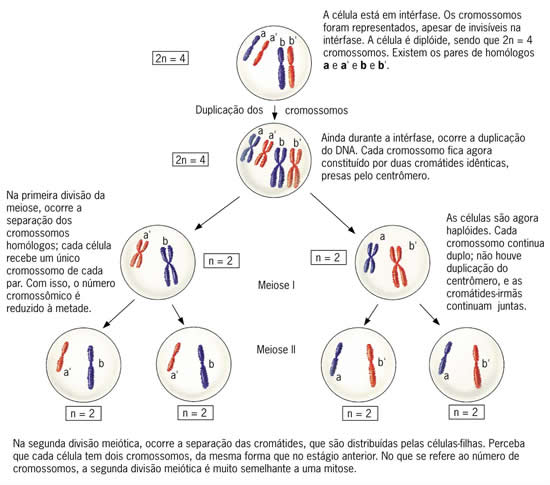
Meiose
Diferentemente da mitose, em que uma célula diplóide, por exemplo, se divide formando duas células também diplóides (divisão equacional), a meiose é um tipo de divisão celular em que uma célula diplóide produz quatro células haplóides, sendo por este motivo uma divisão reducional.Um fato que reforça o caráter reducional da meiose é que, embora compreenda duas etapas sucessivas de divisão celular, os cromossomos só se duplicam uma vez, durante a interfase – período que antecede tanto a mitose como a meiose. No início da interfase, os filamentos de cromatina não estão duplicados. Posteriormente, ainda nesta fase, ocorre a duplicação, ficando cada cromossomo com duas cromátides.
As várias fases da meiose
A redução do número cromossômico da célula é importante fator para a conservação do lote cromossômico das espécies, pois como a meiose formam-se gametas com a metade do lote cromossômico. Quando da fecundação, ou seja, do encontro de dois gametas, o número de cromossomos da espécie se restabelece.
Podemos estudar a meiose em duas etapas, separadas por um curto intervalo, chamado intercinese. Em cada etapa, encontramos as fases estudadas na mitose, ou seja, prófase, metáfase, anáfase e telófase.
Vamos supor uma célula 2n = 2 e estudar os eventos principais da meiose nessa célula.
Meiose I (Primeira Divisão Meiótica)
Prófase I – É a etapa mais marcante da meiose. Nela ocorre o pareamento dos cromossomos homólogos e pode acontecer um fenômeno conhecido como crossing-over (também chamado de permuta)
Como a prófase I é longa, há uma seqüência de eventos que, para efeito de estudo, pode ser dividida nas seguintes etapas:
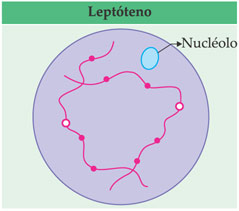
Inicia-se a espiralação cromossômica. É a fase de leptóteno (leptós = fino), em que os filamentos cromossômicos são finos, pouco visíveis e já constituídos cada um por duas cromátides.
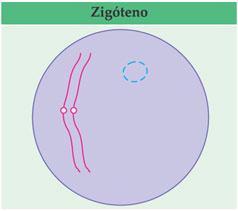
Começa a atração e o pareamento dos cromossomos homólogos; é um pareamento ponto por ponto conhecido como sinapse (o prefixo sin provém do grego e significa união). Essa é a fase dezigóteno (zygós = par).
A espiralação progrediu: agora, são bem visíveis as duas cromátides de cada homólogo pareado; como existem, então, quatro cromátides, o conjunto forma uma tétrade ou par bivalente. Essa é a fase de paquíteno (pakhús = espesso).
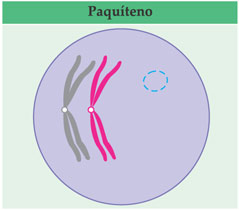
Ocorrem quebras casuais nas cromátides e uma troca de pedaços entre as cromátides homólogas, fenômeno conhecido como crossing-over (ou permuta). Em seguida, os homólogos se afastam e evidenciam-se entre eles algumas regiões que estão ainda em contato. Essas regiões são conhecidas como quiasmas (qui corresponde à letra “x” em grego). Os quiasmas representam as regiões em que houve as trocas de pedaços. Essa fase da prófase I é o diplóteno (diplós = duplo).
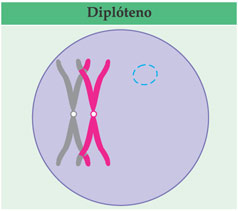
Os pares de cromátides fastam-se um pouco mais e os quiasmas parecem “escorregar” para as extremidades; a espiralação dos cromossomos aumenta. è a última fase da prófase I, conhecida por diacinese (dia = através; kinesis = movimento).
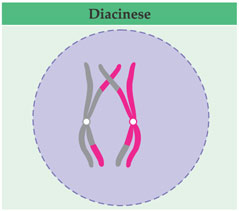
Enquanto acontecem esses eventos, os centríolos, que vieram duplicado da interfase, migram para os pólos opostos e organizam o fuso de divisão; os nucléolos desaparecem; a carioteca se desfaz após o término da prófase I, prenunciando a ocorrência da metáfase I.
Metáfase I – os cromossomos homólogos pareados se dispõem na região mediana da célula; cada cromossomo está preso a fibras de um só pólo.
Anáfase I – o encurtamento das fibras do fuso separa os cromossomos homólogos, que são conduzidos para pólos opostos da célula, não há separação das cromátides-irmãs. Quando os cromossomos atingem os pólos, ocorre sua desespiralação, embora não obrigatória, mesmo porque a segunda etapa da meiose vem a seguir. Às vezes, nem mesmo a carioteca se reconstitui.
Telófase I – no final desta fase, ocorre a citocinese, separando as duas células-filhas haplóides. Segue-se um curto intervalo a intercinese, que procede a prófase II.
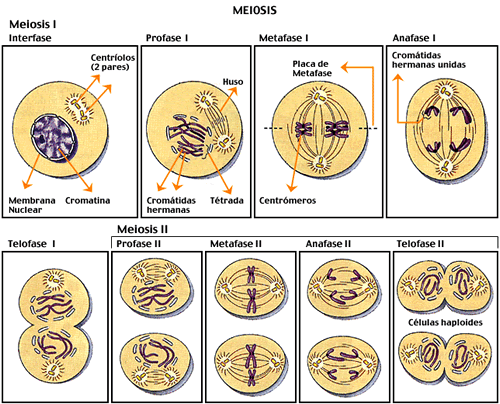
Meiose II (segunda divisão meiótica)
Prófase II – cada uma das duas células-filhas tem apenas um lote de cromossomos duplicados. Nesta fase os centríolos duplicam novamente e as células em que houve formação da carioteca, esta começa a se desintegrar.
Metáfase II - como na mitose, os cromossomos prendem-se pelo centrômero às fibras do fuso, que partem de ambos os pólos.
Anáfase II – Ocorre duplicação dos centrômeros, só agora as cromátides-irmãs separam-se (lembrando a mitose).
Telófase II e citocinese – com o término da telófase II reorganizam-se os núcleos. A citocinese separa as quatro células-filhas haplóides, isto é, sem cromossomos homólogos e com a metade do número de cromossomos em relação à célula que iniciou a meiose.
Variabilidade: Entendendo o crossing-over
A principal conseqüência da meiose, sem dúvida, é o surgimento da diversidade entre os indivíduos que são produzidos na reprodução sexuada da espécie.
A relação existente entre meiose e variabilidade é baseada principalmente na ocorrência de crossing-over.
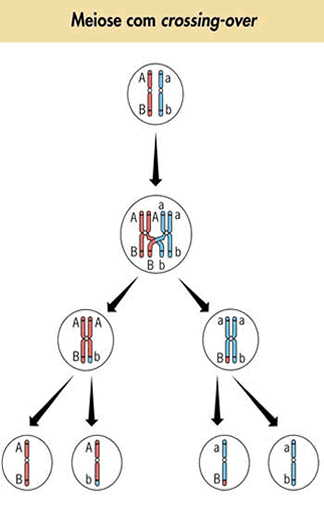
O crossing é um fenômeno que envolve cromátides homólogas. Consiste na quebra dessas cromátides em certos pontos, seguida de uma troca de pedaços correspondentes entre elas.
As trocas provocam o surgimento de novas seqüências de genes ao longo dos cromossomos. Assim, se em um cromossomo existem vários genes combinados segundo uma certa seqüência, após a ocorrência do crossing a combinação pode não ser mais a mesma. Então, quando se pensa no crossing, é comum analisar o que aconteceria, por exemplo, quanto à combinação entre os genes alelos A e a e Be b no par de homólogos ilustrados na figura.
Nessa combinação o gene A e B encontram-se em um mesmo cromossomo, enquanto ae b estão no cromossomo homólogo. Se a distância de A e B for considerável, é grande a chance de ocorrer uma permuta. E, se tal acontecer, uma nova combinação gênica poderá surgir.
As combinações Ab e aB são novas. São recombinações gênicas que contribuem para a geração de maior variabilidade nas células resultantes da meiose. Se pensarmos na existência de três genes ligados em um mesmo cromossomo (A, b e C, por exemplo), as possibilidades de ocorrência de crossings dependerão da distância em que os genes se encontram – caso estejam distantes, a variabilidade produzida será bem maior.
Outro processo que conduz ao surgimento de variabilidade na meiose é a segregação independente dos cromossomos. Imaginando-se que uma célula com dois pares de cromossomos homólogos (A e a, B e b), se divida por meiose, as quatro células resultantes ao final da divisão poderão ter a seguinte constituição cromossômica: (a e b), (a e B), (A e b) e (A e B).
A variabilidade genética existente entre os organismos das diferentes espécies é muito importante para a ocorrência da evolução biológica. Sobre essa variabilidade é que atua a seleção natural, favorecendo a sobrevivência de indivíduos dotados de características genéticas adaptadas ao meio. Quanto maior a variabilidade gerada na meiose, por meio de recombinação gênica permitida pelo crossing-over, maiores as chances para a ação seletiva do meio.
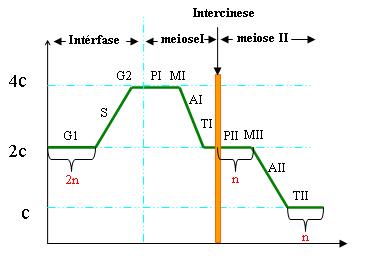
Na meiose a variação da quantidade de DNA pode ser representada como no gráfico ao lado, partindo-se, por exemplo, de uma célula que tenha uma quantidade 2C de DNA em G1.
Gametogênese
Gametogênese é o processo pelo qual os gametas são produzidos nos organismos dotados de reprodução sexuada. Nos animais, a gametogênese acontece nas gônadas, órgãos que também produzem os hormônios sexuais, que determinam as características que diferenciam os machos das fêmeas.O evento fundamental da gametogênese é a meiose, que reduz à metade a quantidade de cromossomos das células, originando células haplóides. Na fecundação, a fusão de dois gametas haplóides reconstitui o número diplóide característico de cada espécie.
Em alguns raros casos, não acontece meiose durante a formação dos gametas. Um exemplo bastante conhecido é o das abelhas: se um óvulo não for fecundado por nenhum espermatozóide, irá se desenvolver por mitoses consecutivas, originando um embrião em que todas as células são haplóides. Esse embrião haplóide formará um indivíduo do sexo masculino. O desenvolvimento de um gameta sem que haja fecundação chama-se partenogênese. Se o óvulo for fecundado, o embrião 2n irá originar uma fêmea.
Em linhas gerais, a gametogênese masculina (ou espermatogênese) e a gametogênese feminina (ovogênese ou ovulogênese) seguem as mesmas etapas.
A Espermatogênese
Processo que ocorre nos testículos, as gônadas masculinas. Secretam a testosterona, hormônio sexual responsável pelo aparecimento das características sexuais masculinas: aparecimento da barba e dos pêlos corporais em maior quantidade, massa muscular mais desenvolvida, timbre grave da voz, etc.As células dos testículos estão organizadas ao redor dos túbulos seminíferos, nos quais os espermatozóides são produzidos. A testosterona é secretada pelas células intersticiais. Ao redor dos túbulos seminíferos, estão as células de Sertoli, responsáveis pela nutrição e pela sustentação das células da linhagem germinativa, ou seja, as que irão gerar os espermatozóides.
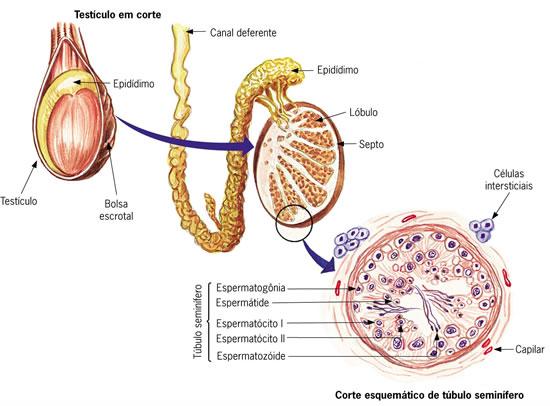
Nos mamíferos, geralmente os testículos ficam fora da cavidade abdominal, em uma bolsa de pele chamada bolsa escrotal. Dessa forma, a temperatura dos testículos permanece aproximadamente 1° C inferior à temperatura corporal, o que é ideal para a espermatogênese.
A espermatogênese divide-se em quatro fases:
Fase de proliferação ou de multiplicação: Tem início durante a vida intra-uterina, antes mesmo do nascimento do menino, e se prolonga praticamente por toda a vida. As células primordiais dos testículos, diplóides, aumentam em quantidade por mitoses consecutivas e formam as espermatogônias .
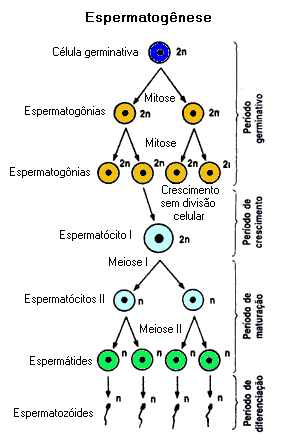
Fase de crescimento: Um pequeno aumento no volume do citoplasma das espermatogônias as converte em espermatócitos de primeira ordem, também chamados espermatócitos primários ou espermatócitos I, também diplóides.
Fase de maturação: Também é rápida, nos machos, e corresponde ao período de ocorrência da meiose. Depois da primeira divisão meiótica, cada espermatócito de primeira ordem origina dois espermatócitos de segunda ordem (espermatócitos secundários ou espermatócitos II). Como resultam da primeira divisão da meiose, já são haplóides, embora possuam cromossomos duplicados. Com a ocorrência da segunda divisão meiótica, os dois espermatócitos de segunda ordem originam quatro espermátides haplóides.
Espermiogênese: É o processo que converte as espermátides em espermatozóides, perdendo quase todo o citoplasma. As vesículas do complexo de Golgi fundem-se, formando o acrossomo, localizado na extremidade anterior dos espermatozóides. O acrossomo contém enzimas que perfuram as membranas do óvulo, na fecundação.
Os centríolos migram para a região imediatamente posterior ao núcleo da espermátide e participam da formação do flagelo, estrutura responsável pela movimentação dos espermatozóides. grande quantidade de mitocôndrias, responsáveis pela respiração celular e pela produção de ATP, concentram-se na região entre a cabeça e o flagelo, conhecida como peça intermediária.
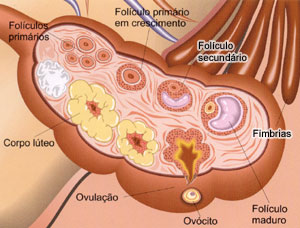
A Ovogênese
Nos ovários, encontram-se agrupamentos celulares chamados folículos ovarianos de Graff, onde estão as células germinativas, que originam os gametas, e as células foliculares, responsáveis pela manutenção das células germinativas e pela produção dos hormônios sexuais femininos.Nas mulheres, apenas um folículo ovariano entra em maturação a cada ciclo menstrual, período compreendido entre duas menstruações consecutivas e que dura, em média, 28 dias. Isso significa que, a cada ciclo, apenas um gameta torna-se maduro e é liberado no sistema reprodutor da mulher.
Os ovários alternam-se na maturação dos seus folículos, ou seja, a cada ciclo menstrual, a liberação de um óvulo, ou ovulação, acontece em um dos dois ovários.
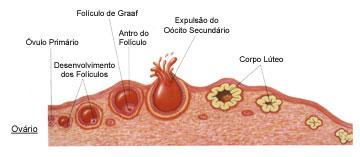
A ovogênese é dividida em três etapas:
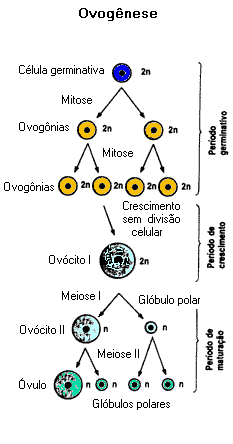
Fase de multiplicação ou de proliferação: É uma fase de mitoses consecutivas, quando as células germinativas aumentam em quantidade e originam ovogônias. Nos fetos femininos humanos, a fase proliferativa termina por volta do final do primeiro trimestre da gestação. Portanto, quando uma menina nasce, já possui em seus ovários cerca de 400 000 folículos de Graff. É uma quantidade limitada, ao contrário dos homens, que produzem espermatogônias durante quase toda a vida.
Fase de crescimento: Logo que são formadas, as ovogônias iniciam a primeira divisão da meiose, interrompida na prófase I. Passam, então, por um notável crescimento, com aumento do citoplasma e grande acumulação de substâncias nutritivas. Esse depósito citoplasmático de nutrientes chama-se vitelo, e é responsável pela nutrição do embrião durante seu desenvolvimento.
Terminada a fase de crescimento, as ovogônias transformam-se em ovócitos primários (ovócitos de primeira ordem ou ovócitos I). Nas mulheres, essa fase perdura até a puberdade, quando a menina inicia a sua maturidade sexual.
Fase de maturação: Dos 400 000 ovócitos primários, apenas 350 ou 400 completarão sua transformação em gametas maduros, um a cada ciclo menstrual. A fase de maturação inicia-se quando a menina alcança a maturidade sexual, por volta de 11 a 15 anos de idade.
Quando o ovócito primário completa a primeira divisão da meiose, interrompida na prófase I, origina duas células. Uma delas não recebe citoplasma e desintegra-se a seguir, na maioria das vezes sem iniciar a segunda divisão da meiose. É o primeiro corpúsculo (ou glóbulo) polar.
A outra célula, grande e rica em vitelo, é o ovócito secundário (ovócito de segunda ordem ou ovócito II). Ao sofrer, a segunda divisão da meiose, origina o segundo corpúsculo polar, que também morre em pouco tempo, e o óvulo, gameta feminino, célula volumosa e cheia de vitelo.
Na gametogênese feminina, a divisão meiótica é desigual porque não reparte igualmente o citoplasma entre as células-filhas. Isso permite que o óvulo formado seja bastante rico em substâncias nutritivas.
Na maioria das fêmeas de mamíferos, a segunda divisão da meiose só acontece caso o gameta seja fecundado. Curiosamente, o verdadeiro gameta dessas fêmeas é o ovócito II, pois é ele que se funde com o espermatozóide.
Fecundação: A volta à Diploidia
Para que surja um novo indivíduo, os gametas fundem-se aos pares, um masculino e outro feminino, que possuem papéis diferentes na formação do descendente. Essa fusão é a fecundação ou fertilização.
Ambos trazem a mesma quantidade haplóide de cromossomos, mas apenas os gametas femininos possuem nutrientes, que alimentam o embrião durante o seu desenvolvimento. Por sua vez, apenas os gametas masculinos são móveis, responsáveis pelo encontro que pode acontecer no meio externo (fecundação externa) ou dentro do corpo da fêmea (fecundação interna). Excetuando-se muitos dos artrópodes, os répteis, as aves e os mamíferos, todos os outros animais possuem fecundação externa, que só acontece em meio aquático.
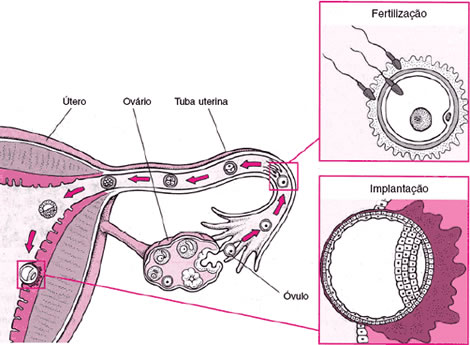
Quando a fecundação é externa, tanto os machos quanto as fêmeas produzem gametas em grande quantidade, para compensar a perda que esse ambiente ocasiona. Muitos gametas são levados pelas águas ou servem de alimentos para outros animais. Nos animais dotados de fecundação interna, as fêmeas produzem apenas um ou alguns gametas por vez, e eles encontram-se protegidos dentro do sistema reprodutor.
Além da membrana plasmática, o óvulo possui outro revestimento mais externo, a membrana vitelínica. Quando um espermatozóide faz contato com a membrana vitelínica, a membrana do acrossomo funde-se à membrana do espermatozóide (reação acrossômica), liberando as enzimas presentes no acrossomo.
As enzimas do acrossomo dissolvem a membrana vitelínica e abrem caminho para a penetração do espermatozóide. Com a fusão da membrana do espermatozóide com a membrana do óvulo, o núcleo do espermatozóide penetra no óvulo. Nesse instante, a membrana do óvulo sofre alterações químicas e elétricas, transformando-se na membrana de fertilização, que impede a penetração de outros espermatozóides.
No interior do óvulo, o núcleo do espermatozóide, agora chamado pró-núcleo masculino, funde-se com o núcleo do óvulo, o pró-núcleo feminino. Cada pró-núcleo traz um lote haplóide de cromossomos, e a fusão resulta em um lote diplóide, o zigoto. Nessa célula, metade dos cromossomos tem origem paterna e metade, origem materna.
Retirado de: http://www.sobiologia.com.br/conteudos/Citologia2/nucleo6.php
Ambos trazem a mesma quantidade haplóide de cromossomos, mas apenas os gametas femininos possuem nutrientes, que alimentam o embrião durante o seu desenvolvimento. Por sua vez, apenas os gametas masculinos são móveis, responsáveis pelo encontro que pode acontecer no meio externo (fecundação externa) ou dentro do corpo da fêmea (fecundação interna). Excetuando-se muitos dos artrópodes, os répteis, as aves e os mamíferos, todos os outros animais possuem fecundação externa, que só acontece em meio aquático.
Quando a fecundação é externa, tanto os machos quanto as fêmeas produzem gametas em grande quantidade, para compensar a perda que esse ambiente ocasiona. Muitos gametas são levados pelas águas ou servem de alimentos para outros animais. Nos animais dotados de fecundação interna, as fêmeas produzem apenas um ou alguns gametas por vez, e eles encontram-se protegidos dentro do sistema reprodutor.
Além da membrana plasmática, o óvulo possui outro revestimento mais externo, a membrana vitelínica. Quando um espermatozóide faz contato com a membrana vitelínica, a membrana do acrossomo funde-se à membrana do espermatozóide (reação acrossômica), liberando as enzimas presentes no acrossomo.
As enzimas do acrossomo dissolvem a membrana vitelínica e abrem caminho para a penetração do espermatozóide. Com a fusão da membrana do espermatozóide com a membrana do óvulo, o núcleo do espermatozóide penetra no óvulo. Nesse instante, a membrana do óvulo sofre alterações químicas e elétricas, transformando-se na membrana de fertilização, que impede a penetração de outros espermatozóides.
No interior do óvulo, o núcleo do espermatozóide, agora chamado pró-núcleo masculino, funde-se com o núcleo do óvulo, o pró-núcleo feminino. Cada pró-núcleo traz um lote haplóide de cromossomos, e a fusão resulta em um lote diplóide, o zigoto. Nessa célula, metade dos cromossomos tem origem paterna e metade, origem materna.
Nenhum comentário:
Postar um comentário